Concept
Fig Wasp and Fig
Intro
A fig is not really a fruit; it is a hollow chamber of flowers turned inward, sealed from the outside world. The only way pollen gets in, and seeds get made, is a wasp barely a couple of millimeters long. A pollen-laden female squeezes through a passage so tight it usually tears off her wings and antennae on the way in, a one-way trip. Inside, she pollinates the flowers, lays her eggs in some of them, and dies. Her offspring hatch, mate in the dark, and the new females load up with pollen from pollen-bearing flowers built for exactly that purpose before tunneling out to find another fig and start over. Each fig species has its own wasp species, and neither can reproduce without the other. A sealed flower that can only be bred by one specific insect, and an insect built and programmed to breed that one flower, is two parts of a single design.
In full
Figs (Ficus, around 750 species) and their pollinating fig wasps (family Agaonidae, such as Blastophaga) form a strict, often one-to-one obligate mutualism: most fig species are pollinated only by their own wasp species, and the wasp can breed only inside that fig. The fig's inflorescence is the syconium, an enclosed receptacle lined internally with tiny flowers, accessible only through a narrow bract-lined ostiole. The female wasp enters through this passage, typically shedding wings and antennae, pollinates the female flowers (actively packing pollen in many lineages, using specialized pollen pockets and combs), oviposits into a subset of flowers that become galls for her larvae, and dies inside. Offspring develop within the syconium; wingless males emerge first, mate with females still in their galls, and chew exit tunnels, after which pollen-laden females leave to repeat the cycle. The system requires a matched, co-dependent set: the sealed syconium and its narrow entry, the wasp's body sized and shaped to enter and its pollen-carrying apparatus, the timed development that synchronizes wasp emergence with the fig's pollen-receptive and pollen-shedding phases, and the fig's regulation of which flowers become seeds versus wasp galls. No single piece functions on its own. A fig with no wasp sets no seed; a wasp with no matching fig has nowhere to breed; an unsynchronized cycle aborts the whole exchange.
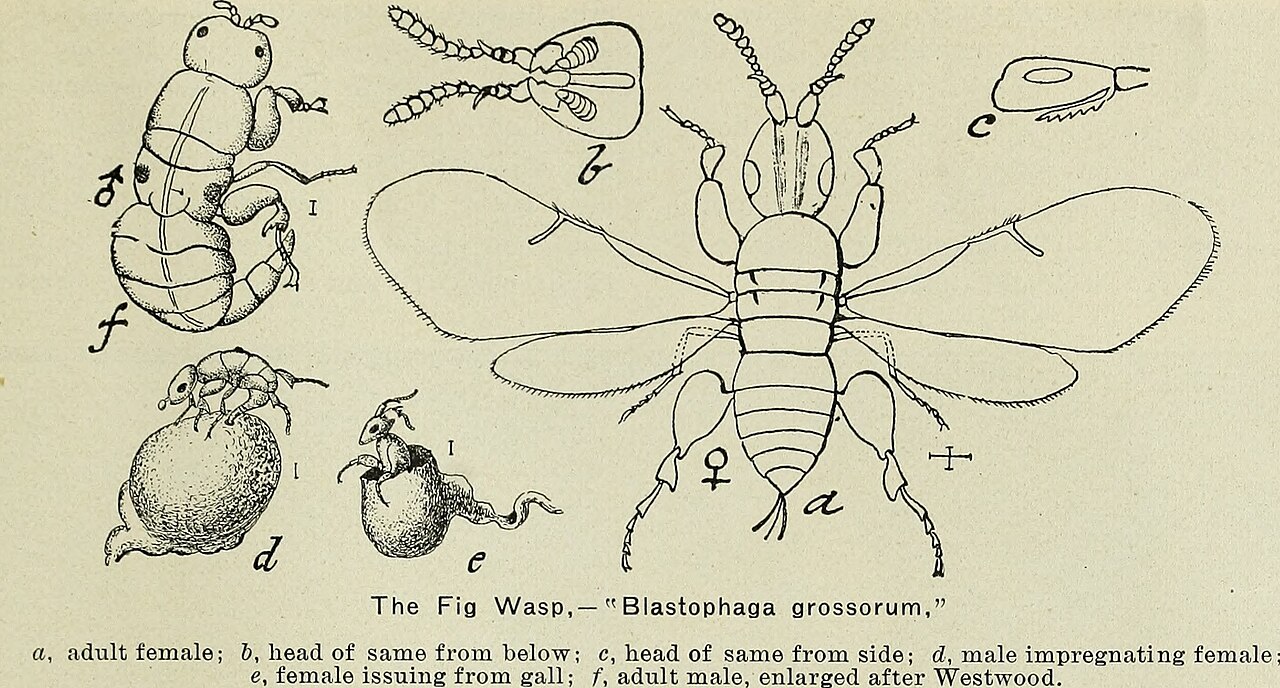
A historical scientific illustration of the fig wasp, Blastophaga grossorum, showing adult female, male, and a female emerging from a gall. Image: public domain, via Wikimedia Commons.
The mechanism
- A sealed chamber. The fig is an enclosed receptacle lined with hundreds of tiny flowers, opening only through a narrow, tightly packed passage.
- A one-way entry. A pollen-bearing female forces through that passage to reach the flowers, usually losing her wings and antennae, committing to the fig for life.
- Pollination and egg-laying. Inside, she pollinates the female flowers and lays eggs in a portion of them, which the fig develops into galls that feed her larvae.
- Mate in the dark. Wingless males hatch first, fertilize the females while they are still in their galls, then cut escape tunnels through the fig wall.
- Loaded for the next fig. New females gather pollen from flowers built to supply it, then leave through the tunnels to find another fig at the right stage and begin again.
Why this points to design
The exchange yields seeds and wasps only when the whole matched set is in place: a sealed syconium with a passage sized for one specific insect, a wasp shaped to enter it and equipped to carry and place pollen, a developmental clock that lines up wasp emergence with the fig's receptive and pollen-shedding phases, and the fig's control over which flowers become seeds and which become wasp nurseries. Pull out any one and nothing works. A fig with no wasp never gets pollinated and sets no seed; a wasp with no matching fig has no chamber to breed in; a passage mistimed against the wasp's life cycle leaves pollen unmoved and eggs unlaid. There is no gradual ramp of separately advantageous steps, because a partly sealed fig with a partly matched wasp and a partly synchronized clock breeds neither party. Two organisms locked together so tightly that each has its own dedicated counterpart, fitted in advance down to body size, timing, and pollen tools, is exactly the prearranged interdependence that points to a designer. See Intelligent Design and Irreducible Complexity.
The evolutionary account, and why it falls short
The standard reply is co-evolution by gradual tightening: ancestral wasps laid eggs in open, ancestral fig-like inflorescences and moved pollen incidentally, and over long spans the receptacle closed up, the wasps specialized, and the two lineages tracked each other into the strict one-to-one bond seen today.
The reply sketches a direction of travel but never supplies the integrated system that needs explaining. Loose egg-laying in an open flower is not the enclosed syconium with a one-insect ostiole, the wing-shedding entry, the synchronized developmental timing, and the active pollen-handling that together define the modern partnership. Naming a route from "open and casual" to "sealed and obligate" is not the same as exhibiting the intermediate fig that could still reproduce while half-sealed, or the wasp that bred in a chamber not yet matched to its body and timing, or the genetic and developmental changes that locked emergence to the fig's pollen phases on both sides at once. The mutual, species-specific, finely timed dependence between a sealed flower and its one wasp is precisely what the gradual story cannot stage and what design accounts for directly.
See also
- Animals That Defy Evolution, the hub this spoke belongs to
- Intelligent Design, the positive design program
- Irreducible Complexity, the matched-parts pattern behind the lock
- Edge of Evolution, the empirical reach of random mutation
- The yucca moth and its yucca plant, another obligate plant-and-insect breeding partnership in this hub
Common questions this page answers
Q: Why is the fig wasp a problem for evolution?
The fig and its wasp depend on each other completely, usually one fig species to one wasp species: the fig is pollinated by nothing else, and the wasp can breed nowhere else. The benefit appears only when the sealed fig, the wasp sized to enter it, the matched pollen-carrying tools, and the synchronized timing are all present together, and a partly built version breeds neither partner. That all-or-nothing, species-specific fit looks engineered rather than assembled by gradual, separately useful steps.
Q: How does fig and fig wasp pollination actually work?
A pollen-bearing female wasp squeezes into the sealed fig through a narrow passage, usually losing her wings and antennae on the way. Inside she pollinates the fig's inward-facing flowers and lays eggs in some of them, then dies; her offspring hatch and mate inside, and the new pollen-loaded females tunnel out to find another fig and repeat the cycle.
Q: Why can't the fig reproduce without its wasp?
The fig's flowers are sealed inside a closed chamber that only the matching wasp can enter, so without that wasp no pollen reaches them and no seeds form. The wasp in turn can develop only inside that fig, and its body size, pollen tools, and life-cycle timing fit that one host. Each is built for the other and useless alone, which is the heart of the design inference here.
Q: Does the wasp harm the fig by laying eggs inside it?
The larvae consume a portion of the fig's flowers, which become galls instead of seeds, but the fig comes out ahead because the wasp guarantees pollination of the rest. The fig also regulates the balance, controlling which flowers become seeds and which become wasp nurseries. The cost is built into a regulated bargain, which keeps the prearranged exchange stable for both.